 By Ken Gilmore
By Ken Gilmore(Editor's Note: Ken is Christadelphian Theist, NOT an Ex-Christadelphian. He does not support or condone the Atheist position of this website and we do not support or condone his Theism. Nevertheless there remains a considerable amount of agreement between us and we consider his work to be of the utmost value)
The following is in five parts with the references given after each part; so keep reading after each section of references.
Why Wrested Scriptures is wrong on evolution - Part 1
Ron Abel’s classic book “Wrested Scriptures”, which looks at parts of the Bible that are often used as proof texts for mainstream theological doctrines such as the Trinity, a supernatural devil and the immortality of the soul has considerable currency in our community. In addition to providing a Christadelphian explanation of these verses, it also strays outside its theological remit and attempts a rebuttal of radiometric dating and evolution. Its attempted rebuttal of these subjects suffers from an uncritical reliance on special creationist material and a flawed understanding of the subject it attempts to criticise. Unfortunately, even though it was written over 40 years ago, and is therefore utterly dated in its attacks on evolution, these attacks still enjoy currency in our community. A detailed critique of the anti-evolutionary arguments of Wrested Scriptures is long overdue.
Over the next few posts, I will take apart the anti-evolution arguments. In so doing, I will be mirroring the section titles of the book.
Abel begins by immediately linking the decline in belief in Biblical inspiration with an acceptance of evolution, and cites the YEC Henry Morris, one of the authors of The Genesis Flood which, given Morris’ poor reputation in the scientific community did not bode well for the rigour of Abels’ arguments.[1] Abel’s attempt to blame evolution for a loss of faith is flat out wrong, as up until the first quarter of the 20th century, theologically conservative Christians had no problem with reconciling evolution and a high view of Scripture. Historian of science David Livingstone observes:
Darwin’s cause in America was championed by the thoroughgoing Congregationalist evangelical Asa Gray, who set himself the task of making sure that Darwin would have “fair play” in the New World. Let us be clear right away that this cannot be dismissed as capitulation to the social pressure of academic peers. To the contrary, Gray had to take on one of the most influential naturalists in America at the time to maintain his viewpoint – none other than Louis Agassiz, a Harvard colleague who vitriolically scorned Darwin’s theory. But Gray was not alone. Many of his countrymen, associates in science and brothers in religion took the same stand. And indeed even those who ultimately remained unimpressed with if not hostile to Darwin were quite prepared to admit that evolution had occurred. It is surely not without significance that Christian botanists, geologists, and biologists – that is to say, those best placed to see with clarity the substance of what Darwin had proposed – believed the evidence supported an evolutionary natural history.[2]
In fact, some of the original authors of The Fundamentals, a collection of books written during the early 20th century to defend orthodox Protestant belief from which the term Fundamentalism is derived, accepted evolution. B.B. Warfield argued that belief in evolution and conservative Christianity were not mutually exclusive, observed that evolution undercut racialist views on polygenism[3] and stated that
“I do not think that there is any general statement in the Bible or any part of the account of creation, either as given in Genesis 1 and 2 or elsewhere alluded to, that need be opposed to evolution.”[4]
James Orr was even more explicit in his support of evolution. Historian Michael Keas, in a 2010 essay on the role R.A. Torrey played in the relationship between evangelicalism and Darwinism in the late 19th and early 20th centuries notes:
In his “Science and Christian Faith” essay, Orr also proposed a resolution to the apparent conflict between biological evolution and the Bible. Significant evidence points to “some form of evolutionary origin of species—that is some genetic connection of higher with lower forms,” but he thought that this change was limited (without specifying how limited). 102 He also argued that God directs the mechanisms of evolution toward purposeful ends. “Evolution,” he concludes, “is coming to be recognized as but a new name for ‘creation’ …”[5]
It is simply wrong to claim that acceptance of evolution is directly correlated to a rejection of belief in an inspired Bible. The fact that Warfield, a theological conservative whose work on inspiration is still influential today saw no essential problem with evolution alone shows the fallacious nature of Abel’s assertion.
After beginning with this demonstrably false claim, Abel attempted to reassure the reader that his or her lack of specialist knowledge would not disqualify them from attempting a definitive rebuttal of evolution:
Many Christians feel an uneasiness about tackling an evolutionist on his own ground. There is usually good reason for this, since one may lack familiarity with the specialized language and data of a particular area of science…Amateur though he may be, there is no monopoly on logic. Every believer should equip himself with sufficient knowledge to clearly formulate the kind of evidence which would be required to reasonably support evolution. Any evidence which may be raised (even if previously unknown) may then be considered in discussion on evolution to avoid the wrangling which can occur over insignificant or even irrelevant considerations. The ground can then be cleared for constructive Biblical teaching to follow.[6]
While a lack of formal scientific education does not disqualify someone from understanding evolutionary biology, or even contributing to the discipline, it is something else altogether for a layperson to think that he is in a position to overturn evolutionary biology if he has not studied or worked in the field. Evolutionary biologist T.R. Gregory points out that:
Over the past 150 years, [Darwin’s initial list of evidence] has been supplemented by countless observations in paleontology, comparative anatomy, developmental biology, molecular biology, and (most recently) comparative genomics, and through direct observations of evolutionary change in both natural and experimental populations. Each of thousands of peer-reviewed articles published every year in scientific journals provides further confirmation (though, as Futuyma notes, “no biologist today would think of publishing a paper on ‘new evidence for evolution’ ... it simply hasn’t been an issue in scientific circles for more than a century”). Conversely, no reliable observation has ever been found to contradict the general notion of common descent. It should come as no surprise, then, that the scientific community at large has accepted evolutionary descent as a historical reality since Darwin’s time and considers it among the most reliably established and fundamentally important facts in all of science. [7]
The folly of Abel’s argument can be readily seen if we replace ‘evolution’ with atomic theory, the germ theory of disease, heliocentrism or any other well-attested scientific theory. Any Christadelphian who had no professional background in those areas would be laughed to scorn if they announced that microbial organisms did not cause disease, the universe revolved around the earth or atoms were mythical. Yet, we do not think it equally risible when a biologically illiterate Christadelphian proudly announces that evolution is false based on nothing more than ‘logic’ and a cursory examination of a popular distillation of the evidence. As Gregory states, evolution is one of the best-attested facts in science, one which is held by the overwhelming majority of professional biologists[8] This is not an appeal to authority, but rather pointing out that if over 99% of professional biologists have examined the evidence in detail (as one would expect given their education and profession) and have concluded that it supports evolution, it is ridiculous to blame the acceptance of evolution on some atheistic conspiracy, particularly given that many of Darwin’s earliest and most ardent supporters were theologically conservative Christians.
What is particularly disturbing about the approach advocated in Wrested Scriptures is the immediate polarising of the discussion created by linking evolution and a decline in Christian belief. By making Christianity and evolution mutually exclusive, this approach is guaranteed to create ex-believers when they examine the evidence for evolution and find it compelling. This alone is reason enough for a takedown of the attempted dismissal of evolution in Wrested Scriptures.
[1] Old Earth Creationist Alan Hayward warned against the growing influence of Henry Morris and John Whitcomb – authors of The Genesis Flood – over 35 years ago. See Hayward A “Flood Geology: A Note of Caution” The Christadelphian (1977) 114:268
[2] Livingstone D.N. “Darwin’s Forgotten Defenders” (Eerdmans 1984) p xi-xii
[3] Warfield B.B. “On The Antiquity and the Unity of the Human Race” Princeton Theological Review (1911) 9:1-25
[4] Cited in Alexander, D (2002) Rebuilding the Matrix: Science and faith in the 21st Century. p. 177.
[5] Keas M “Darwinism,Fundamentalism, and R.A. Torrey” Perspectives on Science and Christian Faith (2010) p 37
[6] Abel R “Wrested Scriptures” Evolution: Preliminary Points
[7] Gregory T.R. “Evolution asFact, Theory and Path” Evo Edu Outreach (2008) 1:46-52
[8] Well over 99% of professional working biologists accept common descent and large scale evolutionary change. See http://phylointelligence.com/dissent.html
In part 2, I examine where Ron Abel's attack on evolution immediately goes astray, and that is in failing to properly define evolution. If you fail to properly define the subject you are attacking, then everything you say loses credibility since you are attacking a parody of the subject.
Abel’s attack on evolution began with a reasonable request to insist on a correct definition of evolution. Unless evolution is properly defined, any discussion will be flawed as what will be attacked is not evolution, but a parody of it. Unfortunately, Abel did just that by failing to define evolution correctly.
He wrote:
1. Insist on a definition of the term evolution. Biology texts use the term in the following three ways:
a) The gradual change in the characteristics of species over the course of time. The Christadelphian need not concern himself with this definition of evolution, since minor changes do occur within species without a change in their essential nature. (E.g. humans today are on the average taller and heavier than humans of a hundred years ago.)
b) 'Horizontal differentiation' - the diversification of a single type into a number of types of creatures with similar characteristics. Again, the Christadelphian need not concern himself with this definition since different types of dogs may have come from one dog type, but they are still dogs.
c) 'Vertical evolution' - the development of existing forms1 from a few primitive forms with increasing complexity and development through the ages. It is this definition which conflicts with the special creative acts of God outlined in Genesis.
Evidence for a) and b) is often mistakenly advanced in support of c).
This is an extremely poor definition of evolution, as it fails to differentiate between evolution as fact (namely the abundant evidence of common descent and large scale evolutionary change) and evolution as theory (the theoretical mechanism proposed to explain how evolution occurred). There was no excuse for Abel’s flawed definition as Darwin himself on at least two occasions was at pains to differentiate between fact and theory. In 1863, four years after the first edition of the Origin was published, Darwin wrote:
Whether the naturalist believes in the views given by Lamarck, or Geoffroy St.-Hilaire, by the author of the Vestiges, by Mr. Wallace and myself, or in any other such view, signifies extremely little in comparison with the admission that species have descended from other species and have not been created immutable; for he who admits this as a great truth has a wide field opened to him for further inquiry.[1]
while in 1871 he wrote in The Descent of Man that:
Some of those who admit the principle of evolution, but reject natural selection, seem to forget, when criticising my book, that I had the above two objects in view; hence if I have erred in giving to natural selection great power, which I am very far from admitting, or in having exaggerated its power, which is in itself probable, I have at least, as I hope, done good service in aiding to overthrow the dogma of separate creations.[2]
Darwin could not have been any clearer in pointing out that when he wrote his book, he had two goals. The first was to show that an evolutionary process had occurred, and to show this he assembled a considerable amount of evidence from biogeography, comparative anatomy and embryology. The second goal was to propose a theoretical explanation for how evolution had occurred, and this was his theory of natural selection.
He was only partly successful in achieving these goals. Within a short time after the publication of his book, the scientific community rapidly accepted that an evolutionary process had occurred. The evidence even then was considered overwhelming. However, he was less successful in convincing the scientific world that natural selection could explain evolution, due in no small part to the lack of a decent theory of heredity. In fact, by the late 19th century, natural selection fell out of favour[3] with alternative theories of evolution such as orthogenesis, mutationism, theistic evolution and Lamarckianism enjoying support until the first half of the 20th century with the creation of the modern evolutionary synthesis, which incorporated natural selection along with insights from genetics, ecology and palaeontology.
However, it needs to be stressed that the fact of evolution was not seriously questioned, as the evidence for it was regarded as overwhelming, even if there was significant debate about the mechanism of evolutionary change. Too many special creationists confuse debate about the theoretical mechanism of evolutionary change with common descent and large-scale evolutionary change, and then erroneously conclude that doubts about the former mean the latter never occurred. This is nonsense, and makes as much sense as concluding that since we do not have a complete theoretical explanation of gravity due to the fact that general relativity does not explain gravity at the quantum level, gravitational lensing, planetary motion and gravitational attraction do not occur. The level of disbelief that a physicist would experience if told this by a layperson is exactly the same experienced by biologists who are confidently told by fundamentalist Christians with no understanding of biology that common descent is false because of problems – real or imagined – with the currently accepted theory of evolution.
The evidence for common descent is considerable, and includes:
- Palaeontology
- Comparative anatomy
- Developmental biology
- Biogeographic distribution of species
- Comparative genomics
It is interesting to note that in the mid-19th century, despite the relative sparseness of the fossil record and the fact that comparative genomics as a discipline was over a century in the future, the evidence for common descent was regarded as compelling. The fossil record and comparative genomics have made a powerful case for common descent irrefutable.
Just the genomic evidence alone is compelling. If common descent was true, then we would expect genetic ‘errors’ such as broken genes, insertion of mobile genetic elements or insertion of retroviral genetic material following viral infection to be inherited by species descending from a common ancestor in which these ‘genetic errors’ first occurred. Conversely, there is no reason if special creation was true for God to deliberately insert the same non-functional genetic element into closely related species in exactly the same place in their respective genomes. What the genomics revolution has shown is that we see many of these ‘genetic errors’ in exactly the same place as predicted by common descent. John Coffin, a leading expert in virology comments on how the pattern of ancient retroviral infection in genomes confirms common descent:
Because the site of integration in the genome, which comprises some three billion base pairs in humans, is essentially random, the presence of an ancient provirus at exactly the same position in different, but related, species cannot occur by chance, but must be a consequence of integration into the DNA of a common ancestor of all the species that contain it. It evolution of retroviruses follows, therefore, that we can infer what viruses were present millions of years ago by examining the distribution of endogenous proviruses in modern species.[4]
As the following video presentation shows, common descent is the only credible explanation for the pattern of endogenous retroviral elements in human and ape genomes. At this point, I need to stress that this is only one of the lines of evidence that confirm common descent.
Having failed to properly define evolution, Abel’s suggested strategy to rebut evolution falls apart. He was attacking a straw man version of evolution, and anyone unfortunate enough to take his strategy to heart and try to take on an informed friend would result in significant embarrassment. There is some value in taking apart the rest of Abel’s flawed attack on evolution if only to show how mistaken contemporary Christadelphians are in offering it as a definitive rebuttal of evolution.
[1] Darwin CR. Origin of species [Letter]. Athenaeum 9 May: 617; 1863.
[2] Darwin C. The descent of man, and selection in relation to sex. London: John Murray; 1871.
[3] This is often referred to as the ‘eclipse of Darwinism’, to use the phrase employed by Julian Huxley.
[4] Coffin JM “Evolution of Retroviruses: Fossils in our DNA” Proceedings of the American Philosophical Society (2004) 148:3, 264-280
Why Wrested Scriptures is wrong on evolution - Part 3
The last post looked at Ron Abel’s failure to properly define evolution by referring to terms such as ‘gradual change in the characteristics of species over time’, ‘horizontal differentiation’ and ‘vertical differentiation.’ This failure to properly differentiate between evolution as fact and evolution as theory meant that Abel’s attack on evolution was fundamentally flawed.
Abel’s failure to properly define evolution can be seen in his attempted rebuttal of the evidence for ‘vertical evolution’, which bears some resemblance to the fact of common descent and large-scale evolutionary change. Abel recognised four of the five main lines of evidence for evolution:
2. It is useful to anticipate that the evidence for 'vertical evolution' is usually taken from four major areas:
- Paleontology - the study of fossil remains of extinct animals and plants, including traces of their existence (e.g. footprints in slate, clay, or coal). It attempts to establish the order in which things lived.
- Morphology and Comparative Anatomy - the study of the comparison of living and dead things for resemblances and differences which might suggest relationships between them and indicate whether one form might have been derived from another.
- Embryology - the study of developing creatures before their birth. Comparisons are made between the adult or developing forms of other creatures.
- Genetics - the study of how the characteristics of parents are transmitted to their offspring, and how variations in these characteristics can arise. It is also concerned experimentally with breeding research in which new variables are produced and specialized.
Missing however is biogeography, which is alluded to only briefly in a footnote. This oversight is damning as the biogeographical distribution of species is one of the strongest lines of evidence for common descent, and shows how rudimentary Abel’s understanding of evolution was. Furthermore, his definition of the evidence for evolution from genetics is not referring to the evidence for common descent from comparative genetics, but rather to the theoretical mechanism behind evolutionary change. This conflation of evolution as fact with evolution as theory stems directly from his flawed definition of evolution.
Before looking at the evidence for common descent from palaeontology, genetics, comparative anatomy, embryology and biogeography, a comment on one of Abel’s footnotes would not go astray. Abel claimed that the main evidence for ‘vertical evolution’ came from four areas, and stated in a footnote that:
Evidence for evolution is sometimes taken from biogeography (the study of geographical distribution of plants and animals) and taxonomy (the science of the classification of organisms). These, however, are not usually the crucial arguments. When arguments of classification are advanced by evolutionists it requires stressing that it is the evolutionist, and not the fossils, which orders the single celled amoeba, first, and next to it places a multicellular hydra, a three-layered worm, an amphioxus, and then a fish, an amphibian, reptile, lower mammal, lower ape, higher ape, and man. The order is a synthetic one. It no more proves that one form of life came from another than the arrangement of books on a shelf from simple to complex proves that the books "evolved". Both simple and complex may have been produced simultaneously. Arguments from the classification assume the very thing which must be proved.
This is nonsense. As I said earlier, the biogeographical distribution of species is one of the strongest lines of evidence for common descent. (More on this later). Furthermore, his assertion that the classification of organisms is entirely arbitrary is false, and betrays a complete ignorance of the principles behind taxonomy.
Take a look at the standard family tree used to classify organisms:
 |
| Source: http://www.talkorigins.org/faqs/comdesc/images/phylo.gif |
This classification system is not arbitrary, but rather consists of a nested hierarchy, that is, groups within groups. Take mammals, the group in which we are classified. Mammals are defined by a number of key features such as a lower jaw consisting of a single bone, three ear bones, and the ability to produce milk. In turn, mammals, birds and reptiles nest in a large group known as amniotes, which are defined as a group of tetrapods that have an egg adapted to land life rather than aquatic life. Most mammals do not lay eggs, but the anatomical structures in the amniote egg have their equivalents in the placenta.
Amniotes, fish and amphibians in turn nest within an even larger group known as vertebrates which are defined as animals that have a vertebral column with a spinal cord dorsal to it, and a gut ventral to it.
Vertebrates, echinoderms (starfish, sea urchins and sea cucumbers) and two other smaller groups of animals themselves fall into an even larger group known as deuterostomes, so called because they share a common feature of embryonic development where the first opening in the early embryo eventually becomes the anus.
Deuterostomes and protostomes (another large group including molluscs and arthropods which are defined by the fact that the first opening in the early embryo becomes the mouth) together form an even higher group called bilaterians which are defined by their possession of bilateral symmetry (clearly defined top, bottom, front and back) as well as in most cases three distinct layers of cells in their embryos.
Bilaterians, comb jellies, sponges and jellyfish in turn nest within the metazoans, or animals which are united by being multicellular, needing to digest food to obtain energy, and lacking rigid cell walls.
The important thing to stress about this family tree is that it is not arbitrary, but driven by objective taxonomic features. Abel was completely wrong when he asserted:
When arguments of classification are advanced by evolutionists it requires stressing that it is the evolutionist, and not the fossils, which orders the single celled amoeba, first, and next to it places a multicellular hydra, a three-layered worm, an amphioxus, and then a fish, an amphibian, reptile, lower mammal, lower ape, higher ape, and man. The order is a synthetic one.
As we can see, this nested hierarchical structure is an objective feature, and not one forced on nature by scientists trying to prove evolution. Furthermore, the only known process that generates a nested hierarchy is descent with modification. This leads to a point overlooked by special creationists. Common descent could easily be falsified by the discovery of many animals that could not be classified in the existing nested hierarchical family tree. Every day, common descent is being tested with the discovery of new animals and plants. Common descent predicts that they will fit within the existing pattern, and to date, every animal and plant discovered fits within this predicted tree.
Conversely, if special creation was true, then there is no reason why animals could be created with features from different nested hierarchies. Feathers make excellent insulation, so there is no reason why the Creator could not have given some mammals feathers. Likewise, given that milk is a good source of food, birds could have been created with the ability to lactate. Likewise, given that whales lactate, fish could have been created with mammary glands. But we don’t see this. Everything in nature falls within a nested hierarchy, exactly as predicted by common descent.
Abel’s complete failure to grasp the reason behind taxonomy betrayed a fundamental ignorance of the subject he set out to criticise. It isn’t a trivial mistake. Animals and plants can be objectively classified in a nested hierarchy, a pattern of groups within groups which is what descent with modification would produce. This family tree is both an objective method of classification, as well as a prediction of the sort of life we should, and should not expect to find.
A final comment would not go astray here. It is also possible to create a family tree based on molecular sequences or gene sequences. Nearly fifty years ago, Linus Pauling and Emile Zuckerkandl predicted that one of the best available proofs of common descent would be the consonance between family trees from morphological and molecular data:
"It will be determined to what extent the phylogenetic tree, as derived from molecular data in complete independence from the results of organismal biology, coincides with the phylogenetic tree constructed on the basis of organismal biology. If the two phylogenetic trees are mostly in agreement with respect to the topology of branching, the best available single proof of the reality of macro-evolution would be furnished. Indeed, only the theory of evolution, combined with the realization that events at any supramolecular level are consistent with molecular events, could reasonably account for such a congruence between lines of evidence obtained independently, namely amino acid sequences of homologous polypeptide chains on the one hand, and the finds of organismal taxonomy and paleontology on the other hand. Besides offering an intellectual satisfaction to some, the advertising of such evidence would of course amount to beating a dead horse. Some beating of dead horses may be ethical, when here and there they display unexpected twitches that look like life."[1]
When the data from molecular and gene sequencing started appearing, the prediction of Pauling and Zuckerkandl was confirmed as phylogenetic trees from molecular data agreed overwhelmingly with the existing phylogenetic tree. This is exactly what common descent would predict.
Conversely, there is no reason for this to be true if special creation was in fact true. Due to redundancy in the genetic code, there are approximately three different ways to code for each amino acid (the building blocks of proteins). This means that for even a moderately large protein such as cytochrome c which has around 104 amino acids, there are 3^104 or 4x10^49 (that's 4 with 49 zeros after it!) possible ways to code for exactly the same protein. What we see is that the coding sequences for cytochrome c cluster together closely in such a way as produce, when graphed, a family tree remarkably consonant with the existing evolutionary family tree.
 |
| Source: Mulligan, P.K. (2008) Proteins, evolution of in AccessScience, ©McGraw-Hill Companies. |
It is hard to ask for a better demonstration of common descent.
References
1 Zuckerkandl E, Pauling L (1965) "Evolutionary Divergence and Convergence in Proteins." in Evolving Genes and Proteins, p. 101.
[2] Common descent would predict that the gene sequences for all animals which have cytochrome c should cluster together in a pattern consistent with descent with modification. Conversely, if special creation was true, there is an unimaginably large coding space for this protein, enough for each animal to have a unique coding sequence separated by so much space as to falsify irrefutably common descent. We do not see this. Either God has been deliberately creating life in such a way as to simulate common descent down to the molecular level (and for no objective reason given that each of those possible 4x10^49 coding sequences produces exactly the same protein), or the remarkable consonance between molecular and morphological phylogenetic trees should be seen as what Zuckerkandl and Pauling stated, the best available proof of common descent.
Ron Abel attempted to show that the fossil record did not support what he called ‘vertical evolution’ by claiming that it needed to show a ‘finely graded sequence’ from simple to complex over evolutionary time. He asserted that the fossil record was discontinuous, showed at best succession rather than descent and concluded by stating that fossils appeared suddenly at the Cambrian. Abel’s argument was a gross misrepresentation of the palaeontological evidence for evolution, and ignored the fact that:
- The insistence on a ‘finely graded sequence’ is a misrepresentation of what we would expect to see in the fossil record based on known mechanisms of speciation
- Evidence for large-scale evolutionary change abounds in the fossil record
- The fossil record is fragmentary because (i) fossilisation occurs only under certain conditions, with some animals fossilising more readily than others (ii) many fossils are destroyed by the movement of the crust over time and (iii) the short amount of time between when fossils are accessible to palaeontologists and when natural processes destroy them
- Fossils do not appear ‘suddenly’ at the Cambrian. Not only is there evidence of complex multicellular life prior to the Cambrian, the appearance took place over several million years.
Its reliance on dated special creationist arguments is painfully obvious.
Evolution? The Fossils say yes.
Abel offered the following strategy for Christadelphians seeking to rebut the evidence for evolution from palaeontology:
- Evidence required - A reasonable argument in support of vertical evolution must show that there is a finely graded sequence from simply to complex over a long period of evolutionary history.
- Evidence lacking
i. At best paleontological enquiry can only show that one form of life came after another, (granting the generous assumption that all inverted orders of fossil deposits can be explained). Present paleontological enquiry does not prove descent.
ii. The fossil record does not show a finely graded sequence from simple to complex. Its record is one of discontinuity. Types spring suddenly in the Cambrian deposits.
He was wrong. The fossil evidence for large-scale evolutionary change is overwhelming. Palaeontologist Steven Gould, in response to special creationist claims that the fossil record showed no evidence of evolutionary transition stated:
Since we proposed punctuated equilibria to explain trends, it is infuriating to be quoted again and again by creationists—whether through design or stupidity, I do not know—as admitting that the fossil record includes no transitional forms. Transitional forms are generally lacking at the species level, but they are abundant between larger groups.[1]
Such transitional forms, as Gould said, abound in the fossil record.[2] One of the better-attested transitions is the evolution of tetrapods from lobe-finned fish, which took place around 380 to 365 million years ago:

Eusthenopteron was a lobe finned fish that lived around 385 million years ago. Acanthostega was a tetrapod, which lives around 365 million years ago. Tiktaalik, a lobe-finned fish that lived around 375 million years ago is a perfect intermediate between tetrapod and fish, with features of both fish and tetrapod. Its front fins have a wrist, while it also has a neck – something fish don’t have. Its mix of fish and tetrapod features have led to the unofficial term ‘fishapod’ to describe this transitional form.
Source: Coyne "Why Evolution is True" (OUP 2009)
Eusthenopteron was a lobe finned fish that lived around 385 million years ago. Acanthostega was a tetrapod, which lives around 365 million years ago. Tiktaalik, a lobe-finned fish that lived around 375 million years ago is a perfect intermediate between tetrapod and fish, with features of both fish and tetrapod. Its front fins have a wrist, while it also has a neck – something fish don’t have. Its mix of fish and tetrapod features have led to the unofficial term ‘fishapod’ to describe this transitional form.
Another well-documented example of large-scale evolutionary change is the evolution of birds from dinosaurs. Birds share many anatomical similarities with the bipedal carnivorous dinosaurs known as theropods. Since the late 20th century, we’ve discovered a considerable number of fossils transitional between birds and theropods, some of which have feathers.

Source: Coyne "Why Evolution is True" (OUP 2009)
The comparison between modern chickens, the transitional fossil Archaeopteryx and the theropod Compsognathus show these similarities. Archaeopteryx had feathers and an opposable great toe, but also had a tail, teeth and reptilian pelvis.

Source: Coyne "Why Evolution is True" (OUP 2009)
One of the more remarkable dinosaur discoveries of recent times is the feathered theropod Sinornithosaurus millenii, which lived around 125 million years ago. Its fossilised remains show clear evidence of filamentous feathers – structures that today are only seen on birds – but whose dinosaur nature is clearly shown by its tail and teeth. Other feathered dinosaurs include Microraptor gui which lived around five million years later than Sinornithosaurus, and had well-formed feathers on its forelimbs and hindlimbs:

Source: Coyne "Why Evolution is True" (OUP 2009)
As with the evolution of tetrapods, where large-scale evolutionary change from fish to tetrapod is clearly seen, the evolution of birds from theropod dinosaurs is fairly clear. We are not looking at succession, but large-scale evolutionary change. This is well seen in the evolution of whales from artiodactyls, hooved mammals. Around fifty million years ago, we see in the fossil record a remarkable transition from hooved mammals to aquatic mammals over a scant ten million year period:

Source: Coyne "Why Evolution is True" (OUP 2009)
This is a cursory overview of a small part of the evidence for large-scale evolutionary change in the fossil record, but it serves to show that contrary to Abel’s assertion, the fossil record shows unarguable evidence of evolution. Geneticist Jerry Coyne notes:
The fossil record teaches us three things. First, it speaks loudly and eloquently of evolution. The record in the rocks confirms several predictions of evolutionary theory: gradual change within lineages, splitting of lineages, and the existence of transitional forms between very different kinds of organisms. There is no getting around this evidence, no waving it away. Evolution happened, and in many cases we see how. [3]
Cambrian Explosion - Slow fuse, not big explosion.
One of the classic creationist claims is that all the major animal groups appeared suddenly at the start of the Cambrian around 541 million years ago, with no evidence of ancestral forms prior to then. Needless to say, Abel’s assertion is wrong. Not only do we have evidence of complex multicellular life before the Cambrian, including ancestors of some of the animal groups that appear in the Cambrian, the appearance did not take place in a geological instant, but over several million years.
Apart from the unique Ediacaran biota, complex metazoan life has been found in the Ediacaran period, and these are usually interpreted as related to modern phyla:
- Vernanimalcula guizhouena: small (< 180 micrometres) organisms believed to be the earliest bilaterians, that is, animals with bilateral symmetry. They have been found [4] in the Doushantuo Formation in southwest China, and are dated to 580-600 million years ago, at least 40 million years before the Ediacaran-Cambrian transition. This interpretation has been challenged by some [5], but if they represent genuine bilaterian fossils [6], they would provide evidence to support the idea that all the modern phyla did not emerge at the start of the Cambrian.
- Kimberella: this is generally accepted as a bilaterian, and is dated at around 558 million years ago, which is about 15 million years before the beginning of the Cambrian. Palaeontologists generally regard it as being related to the molluscs. [7]
- Spriggina: currently considered to be an arthropod, Spriggina [8] bears a passing resemblance to trilobites, and may well be related to them. It has been dated to around 550 million years ago.
 |
| Spriggina floudensi - Source Wikipedia |

Model of Spriggina floundersi.
Image: Benjamin Healley
Source: Museum Victoria
The length of the Cambrian explosion is anything but short. Keith Miller, a palaeontologist at Kansas State University who is also an evangelical Christian notes that if one defines the Cambrian explosion as the length of time from the start of the Cambrian period to the earliest appearance of diverse invertebrate communities in the fossil record, the length of time is around 20 million years.[9] That is a lot of time in which this apparently instantaneous appearance of major phyla appeared. On that point, Miller notes:
However, not all living animal phyla with a fossil record first appear within this time window. The colonial skeleton-bearing bryozoans, for example, are not known from the fossil record until the end of the Cambrian around 491 million years ago. More significantly, several living invertebrate phyla have a fossil record that extends into the late Neoproterozoic before the Cambrian. Sponges have been recognized as early as 580 million years, cnidarians (the group includes jellyfish and anemones) are present among the Ediacaran animals at around 555 million years, and the stem groups...for some other phyla were also likely part of the Ediacaran communities.
Defining the Cambrian “explosion” is not as straightforward as it might seem. Although there was clearly a major burst of evolutionary innovation and diversification in the first 20 million years or so of the Cambrian, this was preceded by an extended period of about 40 million years during which metazoans (multicellular animals) arose and attained critical levels of anatomical complexity. The Ediacaran saw the appearance of organisms with the fundamental features that would characterize the later Cambrian organisms (such as three tissue layers, and bilaterally symmetric bodies with a mouth and anus), as well as the first representatives of modern phyla. The base of the Cambrian is not marked by a sharp dramatic appearance of living phyla without Precambrian roots. It is a subjectively defined point in a continuum. The Cambrian “explosion” appears to have had a “long fuse.” - Emphasis mine
 |
| Source: http://biologos.org/questions/cambrian-explosion |
The ‘Cambrian Explosion’ much touted by creationists, namely the sudden appearance of all major phyla instantaneously at the base of the Cambrian without any evolutionary relationship between each other, is not a problem for science because it never happened, not at least in the way creationists assert.
A finely graded sequence is not what we would expect to see
We’ve seen that contrary to Abel’s assertion, the fossil record shows large scale evolutionary change, not just ‘succession.’ Furthermore, his claim that major classes of animal life appeared suddenly at the start of the Cambrian is false. As Keith Miller puts it, the ‘explosion’ had a long fuse.
What about Abel’s claim that “a reasonable argument in support of vertical evolution must show that there is a finely graded sequence from simply to complex over a long period of evolutionary history”? While there are technical reasons why the fossil record is fragmentary, [10] this is not the reason why we don’t have a ‘finely graded sequence. The reason we don’t is that we have no reason to expect one due to the nature of speciation.
Special creationists often assert that Darwin believed the fossil record should show such a sequence. He did not. In the Origin of Species, Darwin argued against the idea that the transformation of an ancestral population was even and slow (emphasis mine):
But I must here remark that I do not suppose that the process ever goes on so regularly as is represented in the diagram, though in itself made somewhat irregular, nor that it goes on continuously; it is far more probable that each form remains for long periods unaltered, and then again undergoes modification. [11]
"It is a more important consideration ... that the period during which each species underwent modification, though long as measured by years, was probably short in comparison with that during which it remained without undergoing any change." [12]
Darwin also argued against the idea that such a transformation of the ancestral population would involve the entire population or occur over the geographical distribution of this population:
"... natural selection will generally act very slowly, only at long intervals of time, and only on a few of the inhabitants of the same region. I further believe that these slow, intermittent results accord well with what geology tells us of the rate and manner at which the inhabitants of the world have changed." [13]
If Darwin was right, then a fossil record of this process is hardly going to show a ‘finely graded sequence.” Rather, he referred to the process of speciation as being intermittent and irregular, and flatly stated that:
... we have no right to expect to find, in our geological formations, an infinite number of those fine transitional forms, which, on our theory, have connected all the past and present species of the same group into one long and branching chain of life. We ought only to look for a few links, and such assuredly we do find--some more distantly, some more closely, related to each other; and these links, let them be ever so close, if found in different stages of the same formation, would, by many palaeontologists, be ranked as distinct species. [14]
Special creationists are fond of quoting Darwin out of context by touting this quote:
But, as by this theory innumerable transitional forms must have existed, why do we not find them embedded in countless numbers in the crust of the earth?
but failing to continue this quote to provide his answer:
It will be more convenient to discuss this question in the chapter on the Imperfection of the Geological Record; and I will here only state that I believe the answer mainly lies in the record being incomparably less perfect than is generally supposed. The crust of the earth is a vast museum; but the natural collections have been imperfectly made, and only at long intervals of time. [15]
To this one can add Darwin’s comments, which show that he did not expect to see an innumerable series of ‘finely graded transitions’. Ultimately, in order to understand why we don’t see such ‘finely graded transitions’, we need to look at speciation.
Allopatric Speciation and the Fossil Record
Large populations tend to be resistant to evolutionary change as novel genes that arise in this population tend not to be fixed. However, new genes that arise in small populations stand a much better chance of becoming fixed and dramatically changing the gene pool. Examples can be found in small isolated communities where genetic disorders often predominate.
This is the idea behind allopatric speciation, advanced primarily by German evolutionary biologist Ernst Mayr. He argued that the most likely source of new species would be small reproductively isolated populations on the boundaries of larger populations. With gene flow from the larger population to the smaller population eliminated or greatly minimised, the two populations eventually became distinct species which would not interbreed even if the barriers separating them were removed.
The nature of allopatric speciation – something which takes place in small areas over a geological instant – means that we would not expect finely-graded transitions to be seen in the fossil record. Vertebrate palaeontologist Donald Prothero points out that:
If the allopatric speciation model applied to the fossil record, then we should not expect to see speciation in the fossils from the main population. Instead, speciation should occur in small peripherally isolated populations that have little chance of being fossilized. In addition, all of the data from biology showed that this process of speciation typically takes place in tens to hundreds to thousands of years, which is a geological instant as far as paleontologists are concerned. The age difference between two bedding planes is often many thousands of years. Thus we would not expect to see the gradual transitions between species preserved very often; instead, we expect to see new species when they immigrate back in to the main population after their isolation and speciation event. In other words, they would appear suddenly in the fossil record. And once they were established, speciation theory would predict that the main population would remain stable and not change gradually through time but that new species would continually arise on the periphery and migrate back to the homeland. [16]

Source: Prothero "Evolution: What the Fossils Say and Why it Matters"
The diagram above compares what one would expect to see from phyletic gradualism, the ‘finely graded series’ model, and what we generally see in the fossil record.
Sharp eyes readers would have noted the reference to punctuated equilibrium in the diagram above, and concluded that PE is therefore what one would expect from allopatric speciation. Douglas Theobald, in a paper on PE concludes by noting:
Punctuated equilibrium is a valid scientific hypothesis, and when geological strata are complete with good temporal resolution and the fossil record is well-represented, the hypothesis is testable. PE, as construed by Eldredge and Gould, is founded upon the modern allopatric speciation model which lies well within mainstream population genetics. However, PE is not novel, and in large part PE originated with Charles Darwin in The Origin of Species (Darwin credits British paleontologist Hugh Falconer with first proposing that stasis is more predominant in the fossil record than periods of morphological change). Thus, in any meaningful sense of the word, the theory of Punctuated Equilibrium is resolutely "Darwinian." 17
Conclusion
Ron Abel's claim that the fossil record does not support evolution is false. We do not expect to see a 'finely graded sequence' of transitions for two reasons:
- What we know of speciation via the allopatric model states that we would not expect to see those changes, which occur in small, isolated populations over a geological instant, preserved in the fossil record
- The fossil record is fragmentary due to the fact that fossilisation is rare, many animals fossilise poorly and the geologically active nature of the earth means that many fossils are never found.
Contrary to what special creationists assert, Darwin recognised that speciation was intermittent and irregular.
The fossil record does not preserve many transitions at the species level, but at higher levels, they are abundant, and show not succession, but large-scale evolutionary change.
Finally, Abel’s assertions about the Cambrian Explosion are flat-out wrong. It took place over several million years, with evidence of complex multicellular life, including some ancestors of Cambrian organisms present in pre-Cambrian data.
The fossil record shows unarguable evidence of evolution. Believers who use Abel’s suggested strategy in order to rebut evolution will be embarrassed.
References
1. Stephen Jay Gould, Evolution as Fact and Theory, Hen's Teeth and Horse's Toes: Further Reflections in Natural History, New York: W. W. Norton & Company, 1994, p. 260
2. The examples cited appear in Jerry Coyne’s “Why Evolution is True”
3. Jerry Coyne, Why Evolution Is True (2009: Oxford University Press) p 57
4. Chen J et al “Small Bilaterian Fossils from 40-55 million years before the Cambrian” (2004) Science 305:218-22.
5. Bengtson, S.; Budd, G. (2004). “Comment on “small bilaterian fossils from 40 to 55 million years before the Cambrian”. Science 306 (5700): 1291a.
6. Chen, Jun Yuan, Paola Oliveri, Eric Davidson and David J. Bottjer. “Response to Comment on "Small Bilaterian Fossils from 40 to 55 Million Years Before the Cambrian"” (2004) Science 306: 1291
7. Fedonkin, M. A.; Waggoner, B. M. (1997). "The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism". Nature 388 (6645): 868–871
8. Glaessner, Martin F. (1958). “New Fossils from the Base of the Cambrian in South Australia" Transactions of the Royal Society of South Australia 81: 185–188
9. Miller K “The Cambrian ‘Explosion,’ Transitional Forms, and the Tree of Life BioLogos Blog Dec 3 2010
10. Most animals are never fossilised as their bodies are eaten or scavenged before they have a chance of being fossilised. Furthermore, fossilisation is more likely in certain environments (marine) than others (forests) for obvious reasons. Finally, there is the sheer unlikelihood of a fossil being found in the first place before it is eroded or destroyed by natural processes. The fact we have a fossil record in the first place is nothing short of remarkable.
11. Darwin C On the Origin of Species Chapter 4, "Natural Selection," p. 152
12. Darwin, Chapter 10, “On the imperfections of the geological record,” p 428
13. Darwin, Chapter 4, p 140-141
14. Darwin, Chapter 10, p 428
15. Darwin, p 172
16. Donald Prothero, Evolution: What the Fossils Say and Why It Matters (2007: Columbia University Press) p 80
17. Theobald D “All You Need to Know about Punctuated Equilibrium (almost)"
Why Wrested Scriptures is wrong on evolution - Part 5


Atavistic hind limb bones in a humpback whale. Source: TalkOrigins Archive

Why Wrested Scriptures is wrong on evolution - Part 6

Abel’s second attack on what he called ‘vertical evolution’ once again suffered from his failure to properly differentiate between common descent and the theoretical mechanism postulated to explain it. The ‘common designer’ argument fails to take into account the fact that the ‘common designer’ appears constrained to create in a nested hierarchical pattern (groups within groups) which is exactly the process one would expect to see from a process of descent with modification. The fact that the same anatomical mistakes (inverted retina, recurrent laryngeal nerve) are found in groups is consistent with evolution, but makes the common designer look like a serial bungler, repeatedly making the same mistakes in such a way as to simulate common descent. The consonance between molecular and morphological family trees is exactly what one would expect from common descent, but is inexplicable from a special creationist point of view.
His suggested strategy to rebut the morphological, anatomical and embryological data is highlighted below:
Morphological, Anatomical, and Embryological
- Evidence required - It must be shown that x and y are part of an historical sequence of progressive changes, or that x can be made to give birth to y by this or that genetic modification.
- Evidence lacking
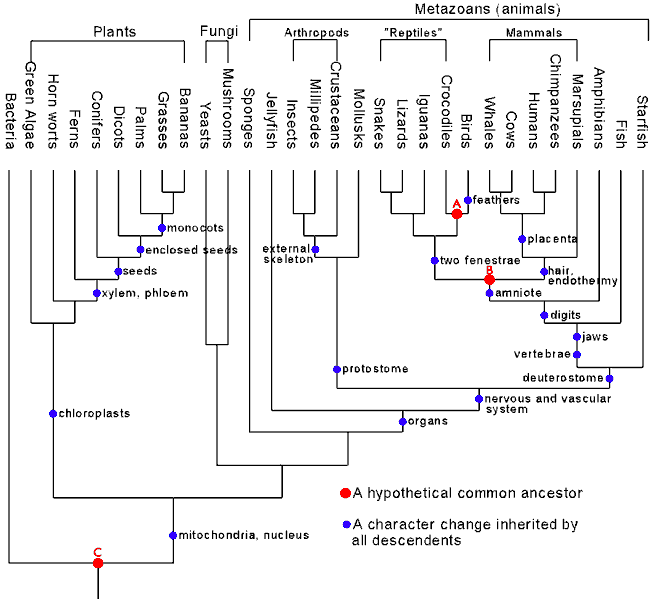
1. Comparing the anatomy of the wing of a bird, the paddle of a whale and the arm of a man, certain similarities of structure can be observed. But resemblances of structure are just as capable of indicating a common designer as ancestry from prior forms.
2. Morphological, anatomical and embryological evidence depend on palaeontological and genetic claims. It depends on whether the fossil record shows a finely graded progression from simple to complex, and whether the study of genetics provides the mechanisms by which creatures would move over time from simple to complex.
I’ve shown that the morphological data does indeed support common descent in an earlier post. Once again, life can be classified objectively in a nested hierarchy, a pattern which is exactly what descent with modification predicts. Asserting that this is evidence of common design is effectively saying that God designed life to look as if it had come about by a process of descent with modification – it is an ad hoc argument employed to argue away evidence which has convinced > 99% of professional biologists that common descent is the better way to explain what we see.
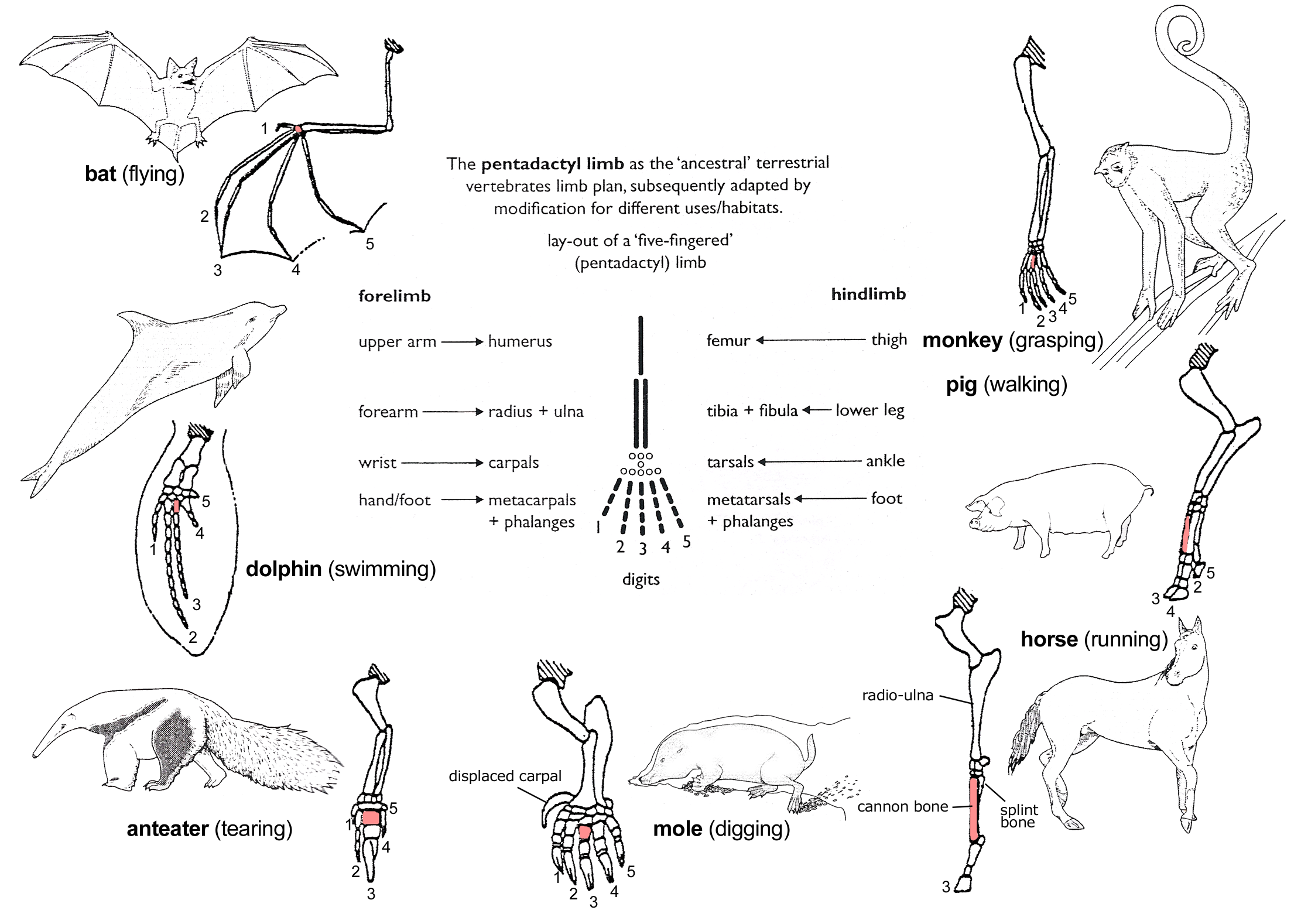
The vertebrate limb is one of the classic examples of how comparative anatomy provides support for common descent. Vertebrates share a common ancestor if common descent is true. Therefore, we'd expect a process of descent with modification to have altered the ancestral vertebrate limb to perform different functions. This is exactly what we see, as the above diagram shows.
Conversely, the common design argument suffers from a number of serious problems:

From a design point of view, this is pointless and wasteful – the nerves supplying the larynx simply need only branch off the vagus nerve as they pass by the larynx and directly supply it.
A knowledge of comparative anatomy and embryology sheds light on this anatomical quirk. In fish, the successive branches of the vagus nerve pass behind the arterial arches running through the gill. Fish do not have a long neck, so this pathway is direct.
Vertebrate embryos initially share this anatomical configuration, which during development is modified. As a result, the path of the recurrent laryngeal nerves loops under arteries such as the right subclavian and the aorta for the right and left RLN, respectively.

A prediction of common descent would be that irrespective of neck length, the RLN would still bypass the larynx, arch under the aorta, and loop up to the larynx. From a design point of view, this would be wasteful and pointless. The extreme case would be that of the giraffe, which would have a couple of metres of wasted nerve. This is exactly what we see:

A recent documentary series highlighted this anatomical evidence for common descent when it featured the dissection of a giraffe:
The vertebrate eye is more evidence for common descent, as it too is an example of shared suboptimal design In the vertebrate eye, the light sensing cells in the retina point away from light. This means light has to pass through blood vessels before reaching the light sensing cells There is a blind spot in vertebrate retinas where blood vessels and nerve fibres pass as they exit and enter the eye. This blind spot would not be needed if the light cells pointed towards light.

Conversely, the common design argument suffers from a number of serious problems:
- There is more than one way to make a wing, grasping limb, flipper, digging limb or running limb, yet we see nothing but variations on the same theme.
- As shown earlier, a comparative anatomical study of life shows nested hierarchies, which means that any designer would be creating in such a way as to simulate common descent. The 'common design' argument then comes across more as as an untestable way to explain away the data
- Shared sub-optimal design is something we'd expect with a blind process such as evolution where initial design errors are inherited by descendant species. An intelligent designer not only can start from a clean sheet, but would not constantly make the same mistakes in the same group.

From a design point of view, this is pointless and wasteful – the nerves supplying the larynx simply need only branch off the vagus nerve as they pass by the larynx and directly supply it.
A knowledge of comparative anatomy and embryology sheds light on this anatomical quirk. In fish, the successive branches of the vagus nerve pass behind the arterial arches running through the gill. Fish do not have a long neck, so this pathway is direct.
Vertebrate embryos initially share this anatomical configuration, which during development is modified. As a result, the path of the recurrent laryngeal nerves loops under arteries such as the right subclavian and the aorta for the right and left RLN, respectively.
A prediction of common descent would be that irrespective of neck length, the RLN would still bypass the larynx, arch under the aorta, and loop up to the larynx. From a design point of view, this would be wasteful and pointless. The extreme case would be that of the giraffe, which would have a couple of metres of wasted nerve. This is exactly what we see:

A recent documentary series highlighted this anatomical evidence for common descent when it featured the dissection of a giraffe:
The vertebrate eye is more evidence for common descent, as it too is an example of shared suboptimal design In the vertebrate eye, the light sensing cells in the retina point away from light. This means light has to pass through blood vessels before reaching the light sensing cells There is a blind spot in vertebrate retinas where blood vessels and nerve fibres pass as they exit and enter the eye. This blind spot would not be needed if the light cells pointed towards light.
This is a design flaw which results in an increased susceptibility towards eye disease. Neurologist Steve Novella notes:
Strangely enough, animals exist with a 'proper' retinal design, where the photoreceptors are in front of the nerve fibres and blood vessels and point towards the light. They are the cephalopods:

The cephalopod eye design is more sensible as the light cells point towards the light, with blood vessels and nerve fibres behind the light sensing layer. The retina is firmly attached to the eye, reducing the likelihood of retinal detachment considerably. Likewise, macular degeneration, and diabetic retinopathy would not be a problem if our eye employed this design. From a special creationist perspective, the vertebrate eye is inexplicable as it is markedly inferior to the cephalopod eye. From the perspective of our evolutionary history, it makes perfect sense.
Embryology
Developmental biologists have recognised for quite some time that early in their development, vertebrate embryos resemble each other closely:

This is known as the phylotypic stage, and poses difficult questions for special creationists. One of them focuses on the shared, counterintuitive way in which the vertebrate face is formed. Structures in the embryonic vertebral arches eventually become facial features.


In all vertebrates, arches 1 and 2 always form the mandible and hyoid bone, respectively. The 3rd and subsequent arches form different structures:
A striking example of how embryology provides powerful evidence for common descent comes from the location of the anatomical structures involved in calcium metabolism. Gills in fish are not only responsible for gas exchange, but in regulation of electrolyte balance. The gills contain sensors and ion pumps that regulate serum calcium levels, making sure that the levels remain at an optimum level.

In humans, we don't have gills, but rather have glands in the neck - the parathyroids and thyroid - to regulate calcium levels. If calcium levels drop too low, then the parathyroid glands signal the kidneys to retain calcium and the bones to release calcium into the blood. Conversely, high calcium levels signal the thyroid which in turn reduces calcium release from bones.
Now, if common descent was true, we would hypothesise that the calcium regulating areas in fish and humans should have the same embryonic origin, as they would have inherited this from the remote common ancestor of fish and humans. We know that in fish, calcium regulation takes place in the gills, which have their embryonic origin in the third and subsequent pharyngeal arches. In humans, the third and subsequent pharyngeal arches form structures in the neck, which is where we find the thyroid and parathyroid glands. I need to stress that there is absolutely no need for these glands to be in the neck. In fact, patients who have their entire thyroids surgically removed due to disease will have their parathyroid glands reimplanted in other anatomical locations.

Impressive as this is, one can go further. In order to confirm whether the parathyroid glands have their embryonic origin in the pharyngeal arches, just as the fish gill sensors have their origin in the pharyngeal arches, we can use molecular biology to test their origin. A molecular marker unique to parathyroids is found in the 3rd and 4th pharyngeal arches in tetrapods. The same marker is found in the pharyngeal arches of fish, even though they don’t have parathyroid glands.
We can test this idea by creating fish that lack the ability to synthesise this marker. These genetically modified fish embryos fail to grow gills. This evidence from anatomy, physiology, molecular biology and developmental biology provide strong evidence supporting the common ancestry of modern fish and modern tetrapods. The common design argument falls apart, as one is entitled to ask why God creates vertebrates in such a way as to consistently simulate common descent.
An earful of jaw - the evolution of the mammalian ear
Perhaps the most spectacular demonstration of common descent from embryology is the developmental biology of the ear and jaw in reptiles and mammals. Mammals have three middle ear bones and one jaw bone, while reptiles have more than one bone in their jaw but only a single middle ear bone.

As both are believed to have a common ancestor, common descent would predict that two bones in the common ancestor which in reptiles form the jaw would in mammals move to form the middle ear.
This is exactly what we see in reptile and mammal embryology. The same two embryonic structures that in reptiles form part of the jaw form two of the middle ear bones in mammals.
When we look at the embryology of the human ear in detail, we see:

This developmental pathway makes sense in the light of common descent and the ear’s evolutionary origin. Once again, the special creationist argument is to simply claim 'common design', without bothering to explain why this 'design' constantly mimics what we'd as a result of common descent. Parsimony favours common descent.
Vestigial structures and atavisms
Vestigial traits are structures which served a function in the ancestor of a species, but either have lost their original function, or have been recruited for other uses. They abound in nature, and provide considerable evidence for common descent. Examples include the wings of flightless birds and the remnants of eyes in marsupial moles. Vestigial traits are also completely misunderstood by special creationists who confuse vestigial with useless. This is not the case. Jerry Coyne comments how:
Dewclaws. These are vestigial digits that can be found on the feet of many vertebrates. In animals that walk on their digits such as dogs and cats, they often do not reach the ground.

Piloerection. This reflex occurs either during cold weather or in response to stress. In the latter, it causes animal to appear larger and more intimidating, while in the former it traps an insulating layer of air. In humans, the response serves neither function and is a relic of our animal ancestry.

Coccyx. Otherwise known as the tail bone. During embryogenesis, human embryos grow a tail, which later degenerates into the coccyx. Its only function now is to serve as an attachment for muscles. The genes for making a tail still exist in humans, and examples of functional tails that contain muscle, nerves and are capable of movement exist.

The top diagram compares the lower spine and tail of a lemur with the lower spine of humans, where the corresponding vertebral bodies C1-C4 (not to be confused with the cervical vertebrae) are fused to form the coccyx. The lower image shows the persistence of the embryonic tail in a baby, and shows the presence of distinct vertebral bodies. (Source: Coyne - Why Evolution is True)
Tooth development in baleen whales. Despite not having teeth, baleen whale embryos grow tooth buds in utero, only to break them down again later in embryogenesis. Transitional fossils of whales with both teeth and baleen-related structures exist, so we know baleen whales evolved from toothed whales. Molecular evidence corroborates this fact as modern baleen whales have broken genes that code for enamel-related protein. The presence of broken genes coding for enamel-related protein in whales that do not have teeth, not to mention the embryonic development of tooth buds that are later broken down is very much an example at the gross anatomical and molecular level of a half-built structure. From an evolutionary point of view, it is intelligible, but from the perspective of special creation, it is inexplicable.

Even worse than the backward arrangement of the rods and cones is the fact that the blood vessels that feed the retinal sit on top of the retina — between the light source and the receptive layer. A more logical arrangement would be to have the blood vessels feed the retina from behind, so that they do not get in the way. In healthy eyes, the blood vessels do not cause any perceptible problem (but they are also partly responsible for the blind spot), but they do limit the total amount of light reaching the rods and cones. The real problem is that they are vulnerable to various diseases. [1]Retinal detachment is one complication of having an inverted retina. If the retina was not inverted, the nerve fibres from the photoreceptors would anchor them securely to the layers beneath them. Other problems arising from having an inverted retina stem from having blood vessels on top of the photoreceptor layer, which means that abnormal blood vessel growth (diabetic retinopathy), bleeding (retinal haemorrhage) or inflammatory changes result in reduced vision.
Strangely enough, animals exist with a 'proper' retinal design, where the photoreceptors are in front of the nerve fibres and blood vessels and point towards the light. They are the cephalopods:

The cephalopod eye design is more sensible as the light cells point towards the light, with blood vessels and nerve fibres behind the light sensing layer. The retina is firmly attached to the eye, reducing the likelihood of retinal detachment considerably. Likewise, macular degeneration, and diabetic retinopathy would not be a problem if our eye employed this design. From a special creationist perspective, the vertebrate eye is inexplicable as it is markedly inferior to the cephalopod eye. From the perspective of our evolutionary history, it makes perfect sense.
Embryology
Developmental biologists have recognised for quite some time that early in their development, vertebrate embryos resemble each other closely:
- All have a notochord and dorsal nerve cord
- All have pharyngeal arches
- All have bodies made from repeated muscle blocks or somites
- All have tails
This is known as the phylotypic stage, and poses difficult questions for special creationists. One of them focuses on the shared, counterintuitive way in which the vertebrate face is formed. Structures in the embryonic vertebral arches eventually become facial features.
In all vertebrates, arches 1 and 2 always form the mandible and hyoid bone, respectively. The 3rd and subsequent arches form different structures:
- Fish: the gills
- Humans: throat cartilage, as well as sundry other muscles and glands
A striking example of how embryology provides powerful evidence for common descent comes from the location of the anatomical structures involved in calcium metabolism. Gills in fish are not only responsible for gas exchange, but in regulation of electrolyte balance. The gills contain sensors and ion pumps that regulate serum calcium levels, making sure that the levels remain at an optimum level.
In humans, we don't have gills, but rather have glands in the neck - the parathyroids and thyroid - to regulate calcium levels. If calcium levels drop too low, then the parathyroid glands signal the kidneys to retain calcium and the bones to release calcium into the blood. Conversely, high calcium levels signal the thyroid which in turn reduces calcium release from bones.
Now, if common descent was true, we would hypothesise that the calcium regulating areas in fish and humans should have the same embryonic origin, as they would have inherited this from the remote common ancestor of fish and humans. We know that in fish, calcium regulation takes place in the gills, which have their embryonic origin in the third and subsequent pharyngeal arches. In humans, the third and subsequent pharyngeal arches form structures in the neck, which is where we find the thyroid and parathyroid glands. I need to stress that there is absolutely no need for these glands to be in the neck. In fact, patients who have their entire thyroids surgically removed due to disease will have their parathyroid glands reimplanted in other anatomical locations.
Impressive as this is, one can go further. In order to confirm whether the parathyroid glands have their embryonic origin in the pharyngeal arches, just as the fish gill sensors have their origin in the pharyngeal arches, we can use molecular biology to test their origin. A molecular marker unique to parathyroids is found in the 3rd and 4th pharyngeal arches in tetrapods. The same marker is found in the pharyngeal arches of fish, even though they don’t have parathyroid glands.
We can test this idea by creating fish that lack the ability to synthesise this marker. These genetically modified fish embryos fail to grow gills. This evidence from anatomy, physiology, molecular biology and developmental biology provide strong evidence supporting the common ancestry of modern fish and modern tetrapods. The common design argument falls apart, as one is entitled to ask why God creates vertebrates in such a way as to consistently simulate common descent.
An earful of jaw - the evolution of the mammalian ear
Perhaps the most spectacular demonstration of common descent from embryology is the developmental biology of the ear and jaw in reptiles and mammals. Mammals have three middle ear bones and one jaw bone, while reptiles have more than one bone in their jaw but only a single middle ear bone.

As both are believed to have a common ancestor, common descent would predict that two bones in the common ancestor which in reptiles form the jaw would in mammals move to form the middle ear.
This is exactly what we see in reptile and mammal embryology. The same two embryonic structures that in reptiles form part of the jaw form two of the middle ear bones in mammals.
When we look at the embryology of the human ear in detail, we see:
- Meckel’s cartilage in the embryo is the cartilaginous bar of the mandibular arch.
- The dorsal ends ossify to form the incus and malleus, while the ventral ends ossify at the tip of the jaw near the chin.
- The middle piece of the cartilage vanishes
This developmental pathway makes sense in the light of common descent and the ear’s evolutionary origin. Once again, the special creationist argument is to simply claim 'common design', without bothering to explain why this 'design' constantly mimics what we'd as a result of common descent. Parsimony favours common descent.
Vestigial structures and atavisms
Vestigial traits are structures which served a function in the ancestor of a species, but either have lost their original function, or have been recruited for other uses. They abound in nature, and provide considerable evidence for common descent. Examples include the wings of flightless birds and the remnants of eyes in marsupial moles. Vestigial traits are also completely misunderstood by special creationists who confuse vestigial with useless. This is not the case. Jerry Coyne comments how:
Opponents of evolution always raise the same argument when vestigial traits are cited as evidence for evolution. “The features are not useless,” they say. “They are either useful for something, or we haven’t yet discovered what they’re for.” They claim, in other words, that a trait can’t be vestigial if it still has a function, or a function yet to be found.
But this rejoinder misses the point. Evolutionary theory doesn’t say that vestigial characters have no function. A trait can be vestigial and functional at the same time. It is vestigial not because it’s functionless, but because it no longer performs the function for which it evolved. The wings of an ostrich are useful, but that doesn’t mean that they tell us nothing about evolution. Wouldn’t it be odd if a creator helped an ostrich balance itself by giving it appendages that just happen to look exactly like reduced wings, and which are constructed in exactly the same way as wings used for flying?
Indeed, we expect that ancestral features will evolve new uses: that’s just what happens when evolution builds new traits from old ones. Darwin himself noted that “an organ rendered, during changed habits of life, useless or injurious for one purpose, might easily be modified and used for another purpose.” [2]Vestigial traits abound in nature; some of the better known examples include:
Dewclaws. These are vestigial digits that can be found on the feet of many vertebrates. In animals that walk on their digits such as dogs and cats, they often do not reach the ground.

Piloerection. This reflex occurs either during cold weather or in response to stress. In the latter, it causes animal to appear larger and more intimidating, while in the former it traps an insulating layer of air. In humans, the response serves neither function and is a relic of our animal ancestry.

Coccyx. Otherwise known as the tail bone. During embryogenesis, human embryos grow a tail, which later degenerates into the coccyx. Its only function now is to serve as an attachment for muscles. The genes for making a tail still exist in humans, and examples of functional tails that contain muscle, nerves and are capable of movement exist.
The top diagram compares the lower spine and tail of a lemur with the lower spine of humans, where the corresponding vertebral bodies C1-C4 (not to be confused with the cervical vertebrae) are fused to form the coccyx. The lower image shows the persistence of the embryonic tail in a baby, and shows the presence of distinct vertebral bodies. (Source: Coyne - Why Evolution is True)
Tooth development in baleen whales. Despite not having teeth, baleen whale embryos grow tooth buds in utero, only to break them down again later in embryogenesis. Transitional fossils of whales with both teeth and baleen-related structures exist, so we know baleen whales evolved from toothed whales. Molecular evidence corroborates this fact as modern baleen whales have broken genes that code for enamel-related protein. The presence of broken genes coding for enamel-related protein in whales that do not have teeth, not to mention the embryonic development of tooth buds that are later broken down is very much an example at the gross anatomical and molecular level of a half-built structure. From an evolutionary point of view, it is intelligible, but from the perspective of special creation, it is inexplicable.
Tooth buds in the jaws of baleen whale embryos
Palmaris longus. in humans, this upper limb tendon which lies between the flexor carpi ulnaris and the flexor carpi radialis muscles has no appreciable function, and in fact is absent in between 10-15% of people. It is often harvested as a tendon graft in hand surgery without any functional deficit. It functions in other animals to expose claws – in humans, this function is no longer necessary and this muscle has become vestigial.
Plantaris. this lower limb muscle has minimal functionality and is absent in up to 10% of the population. As with the palmaris longus, it is often harvested for use in reconstruction surgery with no marked functional deficit, as plantarflexion of the foot is carried out mainly by the gastrocnemius and soleus muscles. In primates, it is used for grasping with the feet, but as we are an obligate bipedal species, we have lost the ability to grasp with the feet and no longer need this function. Again, this is very much an example of a vestigial, “half-functional” structure whose existence makes sense in the light of evolution.
Apomixis in flowering plants. this refers to asexual reproduction via seeds, and is found in a number of plant genera including the dandelions. Put simply, these plants have flowers, despite the fact they do not need them for reproduction. Flowers in apomictic plants are very much a vestigial structure.
Toenails in manatees. these aquatic mammals have flippers and no separate digits, but still retain toenails.

Flightless beetles. there are many flightless beetle species such as the Kauai Flightless Stag Beetle which have fused outer wing covers, but still have inner wings present in a vestigial state. If these beetles were designed from scratch, the presence of useless membranous wings under their fused outer wing covers would be regarded as pointless, wasteful design. Recognition of their evolution from beetles that could fly however makes this design flaw understandable.

Atavisms, or 'throwbacks' to use a vernacular term are the presence of ancestral traits in organisms that are not normally expressed. Examples include hind limbs on whales, toes on horses and tails in humans. Their presence is readily explained by common descent. Normally inactive developmental pathways for traits present in ancestral species but not normally expressed in their descendants are for some reason activated, resulting in the appearance of these 'throwbacks':
Plantaris. this lower limb muscle has minimal functionality and is absent in up to 10% of the population. As with the palmaris longus, it is often harvested for use in reconstruction surgery with no marked functional deficit, as plantarflexion of the foot is carried out mainly by the gastrocnemius and soleus muscles. In primates, it is used for grasping with the feet, but as we are an obligate bipedal species, we have lost the ability to grasp with the feet and no longer need this function. Again, this is very much an example of a vestigial, “half-functional” structure whose existence makes sense in the light of evolution.
Apomixis in flowering plants. this refers to asexual reproduction via seeds, and is found in a number of plant genera including the dandelions. Put simply, these plants have flowers, despite the fact they do not need them for reproduction. Flowers in apomictic plants are very much a vestigial structure.
Toenails in manatees. these aquatic mammals have flippers and no separate digits, but still retain toenails.

Flightless beetles. there are many flightless beetle species such as the Kauai Flightless Stag Beetle which have fused outer wing covers, but still have inner wings present in a vestigial state. If these beetles were designed from scratch, the presence of useless membranous wings under their fused outer wing covers would be regarded as pointless, wasteful design. Recognition of their evolution from beetles that could fly however makes this design flaw understandable.

Brachypterous Colophon westwoodi with vestigial wing visible. [image M. Cochrane, Iziko © ]
Atavisms, or 'throwbacks' to use a vernacular term are the presence of ancestral traits in organisms that are not normally expressed. Examples include hind limbs on whales, toes on horses and tails in humans. Their presence is readily explained by common descent. Normally inactive developmental pathways for traits present in ancestral species but not normally expressed in their descendants are for some reason activated, resulting in the appearance of these 'throwbacks':
Atavistic hind limb bones in a humpback whale. Source: TalkOrigins Archive

Hind limbs in a dolphin. Source: TalkOrigins Archive
This is hardly surprising given common descent, but inexplicable from special creation. After all, if each animal was specially created, why include the developmental pathways for traits that are simply not needed, and more importantly, exactly the pattern of traits which would be expected if common descent were true. Eventually, the tired invocation of 'common design' ceased to have any explanatory power and comes across as simply a desperate attempt to explain away an avalanche of evidence for common descent.
References
References
1. Novella S "Suboptimal Optics: Vision Problems as Scars of Evolutionary History" Evo Edu Outreach (2008) 1:493-497
2. Coyne, J, Why Evolution Is True, (2009: OUP) p 62.
2. Coyne, J, Why Evolution Is True, (2009: OUP) p 62.
Why Wrested Scriptures is wrong on evolution - Part 6
One of the main problems with Ron Abel’s attack on evolution in Wrested Scriptures was his complete failure to properly define evolution, or use his terms consistently. Instead of referring to evolution as fact (common descent) and evolution as theory (modern synthetic theory) he used terms such as ‘horizontal differentiation’ and ‘vertical evolution’ which are hardly standard, and result in a conflation of common descent with the theoretical mechanism proposed to explain it.
This is seen clearly in his attack on the genetic evidence for evolution. In his first two parts (palaeontology and morphology/embryology/comparative anatomy), his arguments were loosely attacking the evidence for common descent. In his third part, he changes approach and attacks the theoretical mechanism, rather than remaining consistent and looking at evidence for common descent as provided by genetics. Although Abel wrote his book well before the genomics revolution, if he had done his research properly, not only would he have recognised the difference between common descent and the modern synthetic theory of evolution, he would have recognised that as early as 1965, biologists had considered the possibility of using amino acid sequences to demonstrate common descent:
"It will be determined to what extent the phylogenetic tree, as derived from molecular data in complete independence from the results of organismal biology, coincides with the phylogenetic tree constructed on the basis of organismal biology. If the two phylogenetic trees are mostly in agreement with respect to the topology of branching, the best available single proof of the reality of macro-evolution would be furnished. Indeed, only the theory of evolution, combined with the realization that events at any supramolecular level are consistent with molecular events, could reasonably account for such a congruence between lines of evidence obtained independently, namely amino acid sequences of homologous polypeptide chains on the one hand, and the finds of organismal taxonomy and paleontology on the other hand. Besides offering an intellectual satisfaction to some, the advertising of such evidence would of course amount to beating a dead horse. Some beating of dead horses may be ethical, when here and there they display unexpected twitches that look like life." – Zuckerkandl, E., Pauling L (1965) "Evolutionary Divergence and Convergence in Proteins." in Evolving Genes and Proteins, p. 101.
This is the genetic evidence that Abel should have referred to if he was to keep his argument consistent. As I’ve stated repeatedly, comparative genomics alone makes the case for common descent irrefutable, with the presence of shared identical genetic errors (ERVs, retrotransposons, pseudogenes) at identical places in human and ape genomes confirming human-ape common ancestry. Arguments about plant breeding and Drosophila genetics not only show that he failed to understand the point, but also failed to understand evolutionary genetics.
Abel offered this advice to the Christadelphian trying to refute the genetic evidence for evolution:
- Evidence required - Experimental evidence to prove that chance factors can elaborate the structural complexities of creatures.
- Evidence lacking
- Although it is true that selective breeding has produced many varieties of plants and animals which may, under carefully controlled conditions, breed true to their new type, the fact that almost without exception, such new types would not be viable in nature is very strong presumption that evolution is unlikely to have been much helped by such processes.
- Indications of diversity on the same level of organization (e.g., Drosophila - the fruit fly) do not prove that present organization has proceeded from single-celled creatures to complex placentae.
- Years of labour, and millions of dollars of research have gone into unlocking the secrets of genetic codes. If anything, this would indicate deliberate planning and highly intelligent understanding behind its formation. It remains to be proven that it could occur by chance.
Abel’s failure to properly differentiate between fact and theory means that his attack on the genetic evidence for evolution is dead in the water. Irrespective of whether the modern evolutionary synthesis needs revision or replacement, the facts of common descent and large scale evolutionary change need to be explained either by the MES or any other theory of evolution. Problems – real or imagined – in the MES no more invalidate the fact of common descent than the inability of general relativity to explain gravity at the quantum level makes gravitational attraction suddenly go away.
Arguably the most powerful demonstration [1] of common descent is the presence of ERV inclusions in the same position in the genome of related species. John Coffin, an acknowledged expert in virology notes that:
Because the site of integration in the genome, which comprises some three billion base pairs in humans, is essentially random, the presence of an ancient provirus at exactly the same position in different, but related, species cannot occur by chance, but must be a consequence of integration into the DNA of a common ancestor of all the species that contain it. It evolution of retroviruses follows, therefore, that we can infer what viruses were present millions of years ago by examining the distribution of endogenous proviruses in modern species. [2]
In a frequently cited paper, Coffin and Johnson pointed [3] out how retroviral inclusions could be employed to reconstruct primate phylogenies or evolutionary family trees using the principle that retroviruses, once fixed in the genome of a species will be inherited by its descendants.
Coffin and Johnson used human endogenous retroviruses - most of the HERV families are found in apes and Old World monkeys. The HERVs used in their study are:
Coffin and Johnson used human endogenous retroviruses - most of the HERV families are found in apes and Old World monkeys. The HERVs used in their study are:
the result of integration events that took place between 5 and 50 million years ago, as indicated by the distribution of specific proviruses at the same integration sites (or loci) among related species. The evolution of primates has been the subject of intense study for well over a century, providing a well established phylogenetic consensus with which to compare and evaluate the performance of ERVs as phylogenetic markers.[4]
What Coffin and Johnson are pointing out towards the end is that we have a fairly reliable evolutionary family tree based on physical characteristics which they used to see how well family trees constructed using ERV data compared with the consensus tree.
Although the technical aspects of constructing such molecular trees are sophisticated, the basic principle behind it is fairly straightforward. An ERV proviral sequence that is selectively neutral will eventually accumulate mutations. If two species share a recent common ancestor, then the ERV inclusions they share will differ by only a small number of mutations, while species that share a remote ancestor will have their common ERV inclusion differing by significantly more mutations. Using sophisticated statistical tools, a family tree can be constructed from the molecular data.
ERV inclusions as Coffin and Johnson point out [5] have three sources of information that can be used to construct phylogenetic trees:
Although the technical aspects of constructing such molecular trees are sophisticated, the basic principle behind it is fairly straightforward. An ERV proviral sequence that is selectively neutral will eventually accumulate mutations. If two species share a recent common ancestor, then the ERV inclusions they share will differ by only a small number of mutations, while species that share a remote ancestor will have their common ERV inclusion differing by significantly more mutations. Using sophisticated statistical tools, a family tree can be constructed from the molecular data.
ERV inclusions as Coffin and Johnson point out [5] have three sources of information that can be used to construct phylogenetic trees:
- The distribution of ERV inclusions among related species
- Accumulated mutations in proviral sequences, which allow an estimate of genetic distance
- Sequence divergence between the LTRs at each end of the ERV inclusion, which is a source of information unique to endogenous retroviruses.
With respect to the first point, both the huge size of the vertebrate genome and the random nature of retroviral integration, it is highly unlikely that one will find multiple ERV inclusions at the same location. As the authors point out:
Therefore, an ERV locus shared by two or more species is descended from a single integration event and is proof that the species share a common ancestor into whose germ line the original integration took place. Furthermore, integrated proviruses are extremely stable: there is no mechanism for removing proviruses precisely from the genome, without leaving behind a solo LTR or deleting chromosomal DNA. The distribution of an ERV among related species also reflects the age of the provirus: older loci are found among widely divergent species, whereas younger proviruses are limited to more closely related species. [6]
The second point is fairly straightforward, as it is similar to the principle underlying other sequence-based phylogenetic analytical methods. As proviral sequences are selectively neutral, they will tend to accumulate mutations at the rate of their occurrence. Two species that have only diverged recently will only differ by a small number of mutations at their common proviral sequence, while those that diverged in the remote past will differ by a larger number of mutations.
The final point - unique to ERVs - is the sequence divergence between the LTRs at either end of the proviral sequence. Of importance here is the fact that as a consequence of reverse transcription, both LTRs will be identical at the time of integration. Johnson and Coffin again:
The final point - unique to ERVs - is the sequence divergence between the LTRs at either end of the proviral sequence. Of importance here is the fact that as a consequence of reverse transcription, both LTRs will be identical at the time of integration. Johnson and Coffin again:
Furthermore, both clusters are predicted to have similar branching patterns as determined by the phylogenetic history of the host species, with similar branch lengths. Thus, each tree displays two estimates of host phylogeny, both of which are derived from the evolution of an initially identical sequence. As we shall see, deviation of actual trees from this prediction provides a powerful means of testing the assumptions and detecting events other than neutral accumulation of mutations in the evolutionary history of a species.[7]
What did Johnson and Coffin find? Lets look at the distribution of the ERVs analysed. In their words:
Three of the loci, HERV-KC4, HERV-KHML6.17, and RTVL-Ia, were detectable in the genomes of OWMs and hominoids, but not New World monkeys, and therefore integrated into the germ line of a common ancestor of the Old World lineages. HERV-K18, RTVL-Ha, and RTVL-Hb were found exclusively in humans, gorillas, chimpanzees, and bonobos, and thus are consistent with a gorilla/chimpanzee/human clade. None of the loci was detected in New World monkeys. [8]
This data is perfectly explained by common descent. To reiterate an “ERV locus shared by two or more species is descended from a single integration event and is proof that the species share a common ancestor into whose germ line the original integration took place.” Johnson and Coffin found many loci shared by these primate species, some shared only by humans, chimps, bonobos and gorillas, some shared only by old world monkeys and hominoids (humans and great apes). This data is consistent with an evolutionary origin of these species, but impossible to explain by special creation without invoking a credulity-stretching concatenation of coincidence.
Estimates of integration time from data obtained from the 5’-LTR and 3’-LTR sequences were also consistent with predictions from common descent:
Estimates of integration time from data obtained from the 5’-LTR and 3’-LTR sequences were also consistent with predictions from common descent:
To estimate the age of each provirus the human/chimpanzee distances from each tree were used to calibrate the rate of molecular evolution at each locus…The most recent common ancestor of humans and chimpanzees lived approximately 4.5 million years ago…so dividing the distance between the human and chimpanzee sequences (substitutions per site) by this number gives rates ranging from 2.3 to 5.0 x 10-9 substitutions per site per year. These numbers are similar to the estimated rates of evolution for pseudogenes and noncoding regions of mammalian genes…Applying each rate to the divergence between the 5’ and 3’ LTRs of the same locus gives integration times consistent with estimates based on species distribution. [9]
Most of the ERVs analysed produced phylogenetic trees consistent with expectation. Their conclusions:
The HERVs analyzed above include six unlinked loci, representing five unrelated HERV sequence families. Except where noted, these sequences gave trees that were consistent with the well established phylogeny of the old world primates, including OWMs, apes, and humans… Phylogenetic analysis using HERV LTR sequences gives rise to trees with a predictable topology, on which is superimposed the phylogeny of the host taxa, and allows ready detection of conversion events.[10]

Evolutionary family trees constructed from ERV data, showing strong agreement with the existing family trees. (Source: Johnson, Coffin.)
In short - we share ERVs with primates at identical places in our genomes which are readily explained by common descent. Furthermore, this is a powerful tool which allows generation of evolutionary family trees. This is but part of the evidence for 'vertical evolution' from genetics, to use Abel's term consistently, and it is compelling evidence for common descent.
Genetic evidence for speciation.
Abel’s insistence on “[e]xperimental evidence to prove that chance factors can elaborate the structural complexities of creatures” repeats the old creationist canard that evolution is about chance. That is misleading [11]. While genetic mutations are random, natural selection is very much a deterministic process.
Selection acts on new genetic diversity, which is produced by a number of mechanisms:
- Point mutation: this is the most common form of mutation, and occurs when the nucleotide at a site in the genome is replaced by a different nucleotide. Some mutations are silent; others can result in a faulty gene product, while others can be beneficial.
- Insertion and deletion: these mutations occur when one or more nucleotides are inserted into, or deleted from a gene. Amino acids are encoded by three consecutive nucleotides, so if these insertions are not multiples of three, the result will be a frameshift mutation that will result in a truncated gene.
- Gene duplication: occasionally, genes (or even whole chromosomes) can be copied, resulting in more than one copy. Mutations in the extra copy can result in a decayed, non-functional gene (pseudogene) or it can result in a gene with a different function, which can allow new functionality to evolve.
- Whole Genome duplication: genomic analysis of many species shows that duplication of the whole genome has occurred in the remote past.
- Insertion of transposons: these are a class of mobile genetic elements that copy and paste, or cut and paste themselves from one part of the genome to another part. Retrotransposons copy themselves via an RNA intermediate, while DNA transposons achieve this transposition without the need for an RNA intermediate. Transposition can cause disease, but has also been linked to the evolution of new functionality.
- Integration of proviral sequences into host genomes: retroviruses, unlike DNA viruses encode their genetic material in RNA. They are able to insert a DNA copy of themselves into the host genome. If they insert themselves into the germline, they can be passed onto the next generation. Depending on where they insert themselves, they can alter the expression of nearby genes. Elements of the proviral sequence can be co-opted by the host genome for completely different uses.
- Horizontal gene transfer: the transfer of genetic elements from one species to another is quite common in bacteria, but has been noted in metazoans. The transferring of antibiotic resistance between different bacterial species is one notable example of horizontal gene transfer.
Some of these changes are deleterious, while some are beneficial. Many other are neutral and may convey an advantage to the population should environmental conditions change later. Contrary to the special creationist distortion of modern evolutionary biology, there are a number of ways in which beneficial mutations occur on which natural selection can work.
Furthermore, natural selection acting on genetic novelty is not the only mechanism of evolutionary change. The neutral theory of evolution argues that evolutionary change at the molecular level can occur through random drift of selectively neutral mutants.
Furthermore, natural selection acting on genetic novelty is not the only mechanism of evolutionary change. The neutral theory of evolution argues that evolutionary change at the molecular level can occur through random drift of selectively neutral mutants.
Some mutations are harmful, some are beneficial but others are neutral. Collyer ignores the fact that a mutation that is beneficial in one environment may be positively harmful in another. The literature is replete with examples of beneficial mutations:
- A point mutation in the apolipoprotein A gene results in the creation of a mutant version of apolipoprotein A, apoA-IM. This beneficial mutation has been linked with reduced rates of atherosclerosis in those heterozygous for the apoA-IM mutation. [12] The clinical significance of this mutation has been explored in many studies. [13][14]
- The CCR5 gene codes for a chemokine receptor that, along with the CD4 T cell co-receptor is used by the HIV-1 virus as an entry point. A mutation in the CCR5 gene conveys resistance to HIV infection in people homozygous for the mutation, while HIV-1 infected individuals heterozygous for the mutation have a two to three year delay before they develop AIDS. The particular mutation is a 32 base pair deletion that results in loss of the CCR5 receptor. [15]
- Bacteria have evolved the ability to metabolise nylon breakdown products. This was achieved by a frameshift mutation which created a unique enzyme that gave the bacteria the opportunity to access a never-before utilised food source. [16]
- In 2008, the evolutionary biologist Richard Lenski published a landmark paper on the evolution by a colony of E. coli bacteria of the ability to metabolise citrate under oxic conditions, something that E. coli is not normally able to do. Over the previous twenty years, Lenski had cultured 12 colonies of E. coli, taking samples every 500 generations to provide a “fossil record”. After 31,500 generations, one colony evolved the ability to metabolise citrate in an oxic environment. The two hypotheses to explain this were (1) an extremely rare mutation or (2) a mutation that was contingent on an earlier mutation to evolve the ability to metabolise citrate. Lenski’s analysis favoured the latter, “Our results instead support the hypothesis of historical contingency, in which a genetic background arose that had an increased potential to evolve the Cit_ phenotype.” [17]
- Evolution of vertebral steroid receptors and the endocrine systems associated with them involved duplication of an ancestral steroid receptor gene, and mutation of the duplicate. [18][19]
- Evidence of chromosomal translocation and segmental duplication in Cryptococcus neoformans. [20]
- Genome duplication in yeast as a source of evolutionary novelty. [21]
- Whole genome duplication as a source of genetic variety is not restricted to yeasts. Examination of the genomes of chordates has shown the existence of multiple copies of genes in the same gene family. None of this is remotely controversial; as long ago as 1999, one paper noted:
Duplication of genes and entire genomes are two of the major mechanisms that facilitated the increasing complexity of organisms in the evolution of life. Gene duplications might be responsible for the functional diversification of genes, the creation of gene families and the generally increased genomic, and possibly also phenotypic, complexity. Protostomes, such as Drosophila, and deuterostome ancestors of vertebrates tend to have single copies of genes whereas chordate genomes typically have more genes, often four; the copies belong to the same gene family. [22]
- Proviral elements from ancient retroviral infections have served as the source of genetic novelty. Placental morphogenesis would not be possible without the cooption of an ancient retroviral envelope protein to perform an entirely different task. [23]
- Horizontal gene transfer in bacteria is responsible for more than just the transfer of antibiotic resistance. One example involves the Salmonella PhoP-PhoQ system which senses environmental magnesium ions, allowing the bacterium to tell whether it is inside a host cell. If this is the case, it activates a molecular pathway that permits it to survive inside the cell. The genes that permit Salmonella to do this are not part of its original genome but were obtained by horizontal gene transfer. [24]
- Evolution of complexity has been simulated and occurs in a Darwinian manner. [25]
The creationist claim that mutations are harmful to life and often lethal is simply wrong, and ignores the fact that as we have shown, mutations can be beneficial. More importantly, he overlooks the fact that most mutations are neutral. Lenski’s long term evolutionary experiment highlights the fact that neutral mutations can linger in the genome until another mutation arrives which effects a beneficial phenotypic change. The average human being has around 175 mutations, [26] according to one study so clearly, most of these are not life-threatening, and otherwise we wouldn’t be here! Really serious mutations tend to be purged from the gene pools via purifying selection. Crippling mutations such as those causing Huntingdon’s disease, CADASIL or other genetic disorders are noticeable because of their characteristic phenotype. Neutral mutations are invisible from a phenotypic perspective, so the special creationist simply overlooks their existence.
Likewise, the creationist attacks on the Drosophila experiments demonstrate specific ignorance of what these experiments are aimed to achieve. [27] The authors of a recent review paper on experimental evolution using a Drosophila model point out:
Experimental evolution with laboratory populations is an alternative tool with which to study adaptation. One of the most attractive applications of experimental evolution is to create a replicated set of populations that have been differentiated relative to replicated control populations, using a well-defined selection protocol, and then compare the allele frequencies at various loci for associations between particular types of phenotypic and genetic differentiation. In short, laboratory evolution allows biologists to use strong inference tests of hypotheses concerning phenotypic and genetic responses to selection. In addition, the recent development of cost-effective genomic tools has allowed broad and systematic assays of the molecular foundations of the effects of experimental evolution. [28]
If one remembers that evolution is a change in allele frequencies in a population, then the 'problem' posed by creationists becomes simply an artefact of the straw man they have created. Examples of evolution observed in Drosophila abound:
Stress-resistance characters respond to direct and reverse selection. Laboratory selection for increased resistance to starvation and desiccation produces rapid responses within populations of D. melanogaster, and high heritability estimates have been reported for both traits. Direct selection for increased starvation (Rose system “SO” and SB” lines) and desiccation (Rose system “D” lines) resistance results in a correlated increase in longevity in the absence of acute starvation or desiccation, along with decreased fecundity...Furthermore, some experiments suggest that lines selected for increased desiccation resistance have decreased preadult viability and slower development time than control lines. There appear to be complex trade-offs connecting resource acquisition during larval stages, adult stress resistance, and life history generally. [29]
Pseudocomparative surveys and reverse-selection experiments reveal correlations between stress resistance and life history characters. The mean longevity of Drosophila populations has been demonstrated to change as a result of manipulating the age of reproduction in a population over multiple generations. Furthermore, longer-lived lines (Rose system “O” lines) have generally been found to tolerate starvation and desiccation significantly better than lines with shorter average lifespans (Rose system “B” lines). These pseudocomparative results were interpreted to mean that resistance to starvation and desiccation might be general physiological mechanisms necessary for maintaining health at late ages. [30]
It hardly needs stressing that there is no ‘limit’ to evolutionary change, but a reference to documented speciation [31] in Drosophila should make this clear, and prevent any further creationist attempts to peddle the usual misunderstandings about Drosophila experiments.
The final – and most important – point focuses on the fact that modern evolutionary biology recognises the importance of networks of genes, and that fixating on single beneficial or deleterious mutations is trivialising evolution. The developmental biologist Paul Myers makes this clear:
The final – and most important – point focuses on the fact that modern evolutionary biology recognises the importance of networks of genes, and that fixating on single beneficial or deleterious mutations is trivialising evolution. The developmental biologist Paul Myers makes this clear:
Mutations are the root of biological variation, of course, but we often have a naive view of their consequences. Most mutations are neutral. Even advantageous mutations are subject to laws of chance in their propagation, and a positive selection coefficient does not mean there will be an inexorable march to fixation, where every individual has the allele. This is also true of deleterious mutations: chance often dominates, and unless it is a strongly negative allele, like an embryonic lethal mutation, there's also a chance it can spread through the population.
Stop thinking of mutations as unitary events that either get swiftly culled, because they're deleterious, or get swiftly hauled into prominence by the uplifting crane of natural selection. Mutations are usually negligible changes that get tossed into the stewpot of the gene pool, where they simmer mostly unnoticed and invisible to selection. Look at human faces, for instance: they're all different, and unless you're looking at the extremes of beauty or ugliness, the variations simply don't make much difference. Yet all those different faces really are the result of subtly different combinations of mutant forms of genes.
"Combinations" is the magic word. A single mutation rarely has a significant effect on a feature, but the combination of multiple mutations may have a detectable or even novel effect that can be seen by natural selection. And that's what's going on all the time: the population is a huge reservoir of genetic variation, and what we do when we reproduce is sort and mix and generate new combinations that are then tested in the environment. [32]
It can be quite difficult to get this simple point across – really serious mutations tend to kill in utero. Even then, some deleterious mutations can be beneficial in some environments, as the persistence of sickle cell anaemia in malaria-endemic areas demonstrates.
Thinking in terms of networks of genes helps realise the fallacy behind the “mutations are deleterious therefore evolution can’t occur” fallacy. Myers again:
Thinking in terms of networks of genes helps realise the fallacy behind the “mutations are deleterious therefore evolution can’t occur” fallacy. Myers again:
The curious thing is, though, that the more elaborate the network, the more pieces tangled into the pathway, the smaller the effect of any individual component (in general, of course). What we find over and over again is that many mutations to any one component may have a completely indetectable effect on the output. The system is buffered to produce a reliable yield.
Now do you see what's wrong with the simplistic caricature of evolution at the top of this article? It's superficial; it ignores the richness of real biology; it limits and constrains the potential of evolution unrealistically. The concept of evolution as a change in allele frequencies over time is one small part of the whole of evolutionary processes. You've got to include network theory and gene and environmental interactions to really understand the phenomena. And the cool thing is that all of these perspectives make evolution an even more powerful force.
Abel’s argument is based on a parody of evolutionary theory, as well as an appalling level of ignorance of the primary literature, not to mention the confused definition of evolution with which he worked.
Conclusion
Abel's suggested strategy to help Christadelphian evolution denialists rebut evolution is hopelessly dated, based on a confused understanding of what mainstream biologists mean by evolution, and deeply indebted to special creationist arguments which were recognised as being wrong in the second half of the 20th century. Anyone using Abel's strategy will find that as John Morris (in a 2009 article attacking evolution!) admitted:
The trouble is, in trying to answer an ‘expert’, we can so readily be wrong-footed. Few of us have advanced qualifications in the relevant sciences, and if we fail to hold our own in discussions about fossils, for example, or if we reveal our ignorance of current molecular biology, we shall be deemed to have lost the contest! [33]
The tragedy of course is that if a believer has been primed to think in terms of evolution versus creation, losing an exchange with a knowledgeable opponent will quite likely precipitate a crisis of faith. The sooner tired anti-evolution arguments such as those made in Wrested Scriptures are buried for good, the better for the intellectual health of our community.
References
2. Coffin JM “Evolution of Retroviruses: Fossils in our DNA” Proceedings of the American Philosophical Society (2004) 148:3, 264-280
3. Johnson WE Coffin JM Constructing primate phylogenies from ancient retrovirus sequences Proc. Natl. Acad. Sci. USA (1999) 96:10254-10260
4. ibid p 10254
5. ibid p 10255
6. loc cit
7. Johnson WE Coffin JM op cit p 10255-10256
8. ibid p 10256
9. ibid p 10259
5. ibid p 10255
6. loc cit
7. Johnson WE Coffin JM op cit p 10255-10256
8. ibid p 10256
9. ibid p 10259
10. loc cit
12. Franceschini G, Sirtori CR, Capurso A, et al. A-I Milano apoprotein: decreased high density lipoprotein cholesterol levels with significant lipoprotein modifications and without clinical atherosclerosis in an Italian family. J Clin Invest. 1980;66:892–900.
13. Sirtori C.R. et al “Cardiovascular Status of Carriers of the Apolipoprotein A-IMilano Mutant” Circulation. 2001;103:1949-1954.
14. Shah P.K. et al “High-Dose Recombinant Apolipoprotein A-IMilano Mobilizes Tissue Cholesterol and Rapidly Reduces Plaque Lipid and Macrophage Content in Apolipoprotein E–Deficient Mice” Circulation. 2001;103:3047-3050.
15. Stevens J.C. et al “Dating the origin of the CCR5-Delta32 AIDS-resistance allele by the coalescence of haplotypes.” Am J Hum Genet. 1998 Jun;62(6):1507-15.
16. Ohno S “Birth of a unique enzyme from an alternative reading frame of the preexisting, internally repetitious coding sequence” Proc. Natl. Acad. Sci. USA (1984) 81:2421-2425
17. Blount Z.D., Borland C.Z., Lenski R.E. "Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli" Proc. Natl. Acad. Sci. USA 2008 105:7899-7906
18. Thornton J.W. "Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions" Proc. Natl. Acad. Sci. USA (2001) 98:5671-5676
19. Bridgham J.T., Carroll S.M., Thornton J.W. “Evolution of Hormone-Receptor Complexity by Molecular Exploitation” Science (2006) 312:97-101
20. Fraser J.A. et al "Chromosomal Translocation and Segmental Duplication in Cryptococcus neoformans" Eukaryotic Cell (2005) 4:401-406
21. Kellis M, Birren B.W, Lander E.S. “Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae” Nature (2004) 428:617-624
22. Meyer A, Schartl M "Gene and genome duplications in vertebrates: the one-to-four (-to-eight in fish) rule and the evolution of novel gene functions" Curr Opin Cell Biol 1999, 11:699-704
23. Mi S et al "Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis" Nature (2000) 403:785-789
24. McAdams H.H. Srinivasan B, Arkin A.P "The Evolution of Genetic Regulatory Systems in Bacteria" Nat Rev Genet (2004) 5:169-178
25. Lenski R.E., Ofria C, Pennock R.T., Adami C “The evolutionary origin of complex features” Nature (2003) 423:139-144
26. Nachman, M.W., Crowell, S.L. Estimate of the mutation rate per nucleotide in humans. Genetics 2000 156(1): 297-304.
27. Burke M.K., Rose M.R., “Experimental evolution with Drosophila” Am J Physiol Regul Integr Comp Physiol 296:R1847-R1854, 2009
28. ibid, p R1847
29. ibid, p R1849
30. loc cit.
31. Dobzhansky, T, Pavlovsky, O "An experimentally created incipient species of Drosophila", 1971 Nature 23:289-292.
32. Myers PZ “It’s more than genes, it’s networks and systems.” Pharyngula July 24th 2010. Accessed 27th Oct 2013.
13. Sirtori C.R. et al “Cardiovascular Status of Carriers of the Apolipoprotein A-IMilano Mutant” Circulation. 2001;103:1949-1954.
14. Shah P.K. et al “High-Dose Recombinant Apolipoprotein A-IMilano Mobilizes Tissue Cholesterol and Rapidly Reduces Plaque Lipid and Macrophage Content in Apolipoprotein E–Deficient Mice” Circulation. 2001;103:3047-3050.
15. Stevens J.C. et al “Dating the origin of the CCR5-Delta32 AIDS-resistance allele by the coalescence of haplotypes.” Am J Hum Genet. 1998 Jun;62(6):1507-15.
16. Ohno S “Birth of a unique enzyme from an alternative reading frame of the preexisting, internally repetitious coding sequence” Proc. Natl. Acad. Sci. USA (1984) 81:2421-2425
17. Blount Z.D., Borland C.Z., Lenski R.E. "Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli" Proc. Natl. Acad. Sci. USA 2008 105:7899-7906
18. Thornton J.W. "Evolution of vertebrate steroid receptors from an ancestral estrogen receptor by ligand exploitation and serial genome expansions" Proc. Natl. Acad. Sci. USA (2001) 98:5671-5676
19. Bridgham J.T., Carroll S.M., Thornton J.W. “Evolution of Hormone-Receptor Complexity by Molecular Exploitation” Science (2006) 312:97-101
20. Fraser J.A. et al "Chromosomal Translocation and Segmental Duplication in Cryptococcus neoformans" Eukaryotic Cell (2005) 4:401-406
21. Kellis M, Birren B.W, Lander E.S. “Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae” Nature (2004) 428:617-624
22. Meyer A, Schartl M "Gene and genome duplications in vertebrates: the one-to-four (-to-eight in fish) rule and the evolution of novel gene functions" Curr Opin Cell Biol 1999, 11:699-704
23. Mi S et al "Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis" Nature (2000) 403:785-789
24. McAdams H.H. Srinivasan B, Arkin A.P "The Evolution of Genetic Regulatory Systems in Bacteria" Nat Rev Genet (2004) 5:169-178
25. Lenski R.E., Ofria C, Pennock R.T., Adami C “The evolutionary origin of complex features” Nature (2003) 423:139-144
26. Nachman, M.W., Crowell, S.L. Estimate of the mutation rate per nucleotide in humans. Genetics 2000 156(1): 297-304.
27. Burke M.K., Rose M.R., “Experimental evolution with Drosophila” Am J Physiol Regul Integr Comp Physiol 296:R1847-R1854, 2009
28. ibid, p R1847
29. ibid, p R1849
30. loc cit.
31. Dobzhansky, T, Pavlovsky, O "An experimentally created incipient species of Drosophila", 1971 Nature 23:289-292.
32. Myers PZ “It’s more than genes, it’s networks and systems.” Pharyngula July 24th 2010. Accessed 27th Oct 2013.
No comments:
Post a Comment
Please do not comment as 'Anonymous'. Rather, choose 'Name/URL' and use a fake name. The URL can be left blank. This makes it easier to see who is replying to whom.